
Cosa sono I recettori del gusto
Nella difesa antimicrobica delle vie respiratorie sta emergendo una nuova classe di recettori: è la famiglia dei recettori del gusto amaro, detti “Taste Receptors” (T2R). Si tratta di recettori accoppiati a proteine G, tipicamente espressi nelle papille gustative, la cui funzione è quella di riconoscere i sapori, anche al fine di prevenire l’ingestione di sostanze tossiche, inclusi prodotti batterici o veleni vegetali. Attraverso lo stesso meccanismo che consente di rilevare i cibi dal sapore amaro i recettori T2R espressi nelle vie respiratorie svolgerebbero un ruolo importante nella protezione da sostanze nocive e infezioni batteriche.
Il nostro cibo è, e deve essere, chimico e per riconoscere la sua composizione due dei nostri sensi sono, appunto, sensi chimici: l’olfatto e il gusto. L’olfatto ci permette di identificare alcuni composti, quelli volatili, a distanza, mentre il gusto ha la funzione di analizzare il contenuto di un alimento riconoscendo le sostanze chimiche di cui è costituito, permettendoci di distinguere i cibi ricchi di nutrienti indispensabili per il nostro sostentamento (che ingoiamo) da quelli potenzialmente tossici o avariati (che rifiutiamo).
Cosa si intende per codice del gusto?
Da qui il codice del gusto: gradiamo il dolce (segnale di carboidrati), l’umami (segnale di proteine) e il salato (Na+), mentre non gradiamo l’acido, segnale di frutti non maturi o di fermentazioni, e l’amaro (composti potenzialmente tossici prodotti prevalentemente da piante). Ovviamente questa è una semplificazione, dato che esistono composti dolci molto tossici e composti amari che non lo sono.
Alcuni di noi hanno scritto tempo fa su questa rivista in tema di gusto e a quell’articolo1 rimandiamo, sia per maggiori informazioni circa il codice del gusto, sia perché sempre più convinti della possibilità di educare il gusto nei bambini e dell’importanza di fare ciò al fine di instaurare preferenze alimentari “virtuose” con effetti positivi sulla salute.
L’acqua è davvero insapore?
Oltre ai cinque gusti fondamentali, il sistema del gusto è in grado di rispondere anche alle “bollicine” causate dalla CO2, che possono modulare la percezione dei sapori, riducendo, in particolare, quella del dolce2. Invero le bevande gassate aumentano il consumo di glucosio, diminuendone la percezione, e aumentano così il rischio di un eccessivo introito di calorie specie nel bambino.
Quali sono i recettori del gusto ?
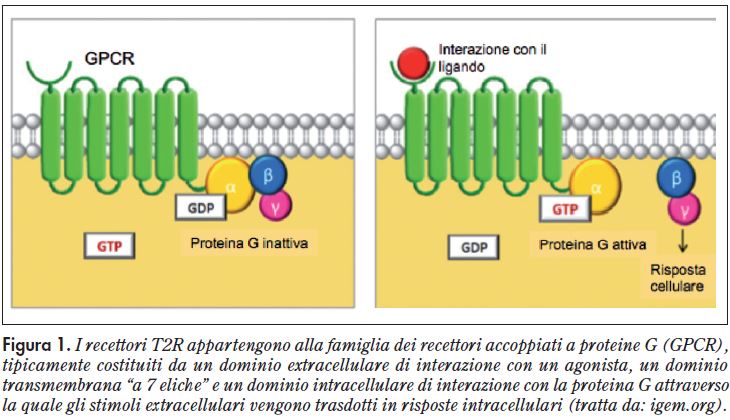
I recettori per il gusto dolce, umami e amaro, appartengono alla classe dei recettori accoppiati a proteine G, generalmente indicati come GPCR (G Protein-Coupled Receptor): sono dei recettori costituiti da 7 eliche che attraversano la membrana cellulare3.
Il legame con la molecola segnale avviene nella parte extracellulare o transmembrana, inducendo un cambiamento conformazionale del GPCR che è in grado di innescare la sua interazione con la vicina proteina G. Quindi, nei recettori accoppiati a proteine G, un segnale chimico è tradotto all’interno di una cellula nel cambiamento di concentrazione di un “secondo messaggero”, generalmente lo ione Ca++ (Figura 1).
Dove si trovano i recettori del gusto?
Recentemente i recettori gustativi appartenenti alla classe dei GPCR, quindi i recettori di dolce, umami e amaro, sono stati identificati anche in altre sedi anatomiche, quali lo stomaco, l’intestino e l’apparato respiratorio4-5.
A cosa servono i recettori del gusto?
La funzione dei recettori dei sapori dolce e umami nello stomaco e nell’intestino è sempre di sensori di nutrienti, influendo sul rilascio di ormoni e/o sulla sintesi di trasportatori di nutrienti.
Ad esempio l’attivazione del recettore del sapore dolce T1R2T1R3 nelle cellule enteroendocrine dell’apparato digerente porta alla produzione di incretine quali il peptide GLP-1 (Glucagon-Like Peptide), che a sua volta favorisce il rilascio di insulina6. Inoltre, l’attivazione dello stesso recettore aumenta anche l’espressione dei trasportatori del glucosio SGLT1 (cotrasportatore attivo di sodio/glucosio) e GLUT2 (trasportatore passivo) nella parte apicale delle cellule dell’epitelio intestinale, aumentando in questo modo l’assorbimento di glucosio.
Perchè a volte ho l’“amaro in bocca”?
Per il gusto amaro sono stati identificati circa 25 recettori funzionali, indicati con T2Rs. Nel cavo orale la loro funzione è il riconoscimento di sostanze potenzialmente tossiche, mentre non sono completamente note le funzioni che assolvono nell’intestino. È stato provato che nella prima parte dell’intestino la loro attivazione porta al rilascio di colecistochinina (CCK), con tutti i suoi effetti, tra i quali la riduzione della motilità intestinale e quindi dello svuotamento gastrico, fenomeni che porterebbero a minore assunzione delle sostanze amare potenzialmente tossiche che hanno attivato i recettori T2Rs)7. Invece nel colon la loro attivazione induce la secrezione di anioni e altri fluidi, con un effetto di “flush out” che favorisce l’eliminazione di eventuali composti tossici amari giunti fino a quella parte dell’intestino.Quali ricettori sono presenti nel naso?
Cellule chemosensoriali "sentinelle" delle respirazione e cellule epiteliali ciliali
Studi funzionali condotti sui topi8 hanno dimostrato che la somministrazione per via nasale di composti amari, come il chinino, produceva rallentamento della velocità di respirazione e apnea, risposte tipicamente evocate sulle terminazioni trigeminali da sostanze a effetto irritante come l’ammoniaca. La via attraverso la quale vengono evocate le risposte trigeminali coinvolge un particolare tipo di cellule sensoriali specializzate che raccordano “il mondo esterno” con il nostro sistema neurovegetativo, le “cellule chemosensoriali solitarie" (SCC) intersperse nell’epitelio della mucosa respiratoria.
A cosa servono le cellule SCC?
Le cellule chemosensoriali sono appunto in grado di tradurre stimoli chimici in riflessi locali. Esse sono infatti dotate di microvilli su cui sono presenti recettori T2R che si affacciano nelle cavità nasali e sono connesse a fibre trigeminali. Attraverso le cellule SCC il sistema di rilevamento dei composti irritanti può rispondere a molecole intrappolate nello strato di muco che copre l’epitelio, stimolando i nervi trigeminali ed evocando riflessi protettivi quali starnuto, tosse, apnea9.
Le cellule SCC appaiono quindi strategicamente situate nell’epitelio respiratorio per monitorare sostanze tossiche presenti nel muco che riveste le superfici respiratorie.
I recettori TAS2R presenti sulle cellule SCC rispondono agli acil-omoserina-lattoni (AHLs) utilizzati come molecole di segnalazione intercellulare dai batteri Gram-negativi, agendo su meccanismi importanti per le difese mucosali10.
Dunque la capacità di intercettare molecole utilizzate dai Gram-negativi come segnali quorum sensing (vedi Box 1, pag. 443) fornisce all’epitelio respiratorio la possibilità di innescare una risposta infiammatoria, prima che i batteri raggiungano una densità di popolazione capace di produrre biofilm patogeni e fattori di virulenza.
Cosa sono le cellule epiteliali?
Le cellule epiteliali ciliate sono dotate di un sistema di difesa dai patogeni dipendente dai recettori T2R
Le cellule ciliate dell’epitelio respiratorio naso-sinusale svolgono un importante ruolo nell’immunità sia innata che adattativa. La clearance mucociliare rappresenta per l’apparato respiratorio la prima strategia innata di difesa nei confronti di patogeni, tossine o sostanze irritanti.
A cosa servono le cellule epiteliali?
L’espressione di recettori T2R nell’epitelio ciliato delle vie respiratorie e specificamente nelle ciglia è stata per la prima volta dimostrata nel 200911: alcuni composti amari quali il chinino, la nicotina, la salicina (che attivano specifici recettori T2R) sono in grado di stimolare la frequenza del battito ciliare, suggerendo quindi che le ciglia sono in grado di agire da “sensori” di composti tossici inalati e attivare una risposta meccanica di difesa.
Dunque è ipotizzabile che in maniera analoga i recettori T2R siano in grado di riconoscere prodotti batterici ed esplicare un’azione protettiva nei confronti di organismi patogeni.
Ciò è stato dimostrato utilizzando colture di cellule ciliate dei seni nasali. In vitro queste cellule sono capaci di rispondere a concentrazioni fisiologiche di AHLs secreti da Pseudomonas aeruginosa inducendo, attraverso un incremento del Ca++ intracellulare, l’attivazione della NO sintetasi e la produzione di ossido nitrico che svolge due ruoli fondamentali nella difesa innata: stimolazione del battito ciliare e quindi della clearance mucociliare e azione battericida diretta12.
Cellule SCC e cellule epiteliali: quale differenza?
Quindi, mentre il ruolo delle cellule SCC sembra essere quello di prevenire l’ulteriore inalazione di sostanze tossiche (attraverso le risposte trigeminali), quello delle cellule ciliate potrebbe consistere nel montare una rapida risposta immunitaria di tipo innato.
Questi risultati sono di grande interesse per il trattamento delle patologie in cui la ridotta efficienza del meccanismo di clearance mucociliare nelle vie respiratorie superiori può risultare in infezioni batteriche ricorrenti, come le rinosinusiti croniche (Chronic Rhinosinusitis- CRS) e apre nuovi spiragli a strategie terapeutiche alternative all’uso (e abuso) di antibiotici, basate sulla stimolazione delle difese innate.
Nell’uomo un ruolo specifico nella risposta agli AHL è stato dimostrato per il recettore TAS2R38 espresso nelle ciglia delle cellule epiteliali delle alte vie respiratorie.
Sostanze amare: una nuova classe di broncodilatatori?
Diverse sostanze amare possono attivare i TAS2Rs espressi nella muscolatura liscia delle vie respiratorie determinando una potente broncodilatazione.
Se cellule muscolari lisce delle vie respiratorie vengono cimentate in vitro con sostanze amare come clorochina o chinino, agonisti di vari recettori T2R, in presenza di broncocostrittori, si registra un effetto miorilassante, molto più efficace rispetto a quello indotto dagli agonisti dei recettori beta2-adrenergici, i più comuni broncodilatatori13.
L’effetto “broncodilatatore” dei composti amari è T2R-dipendente, come dimostra l’evidenza che inibitori farmacologici di molecole importanti nella via di segnalazione dei recettori T2R prevengono l’effetto della clorochina o del chinino.
Questo risultato, piuttosto sorprendente, apre la via all’identificazione di nuovi broncodilatatori, efficaci e sicuri, tra le migliaia di composti amari naturali o sintetici oggi disponibili. Prospettiva questa molto promettente, considerata la prevalenza di patologie respiratorie ostruttive (più di 300 milioni di persone nel mondo) e il ruolo centrale che le disfunzioni della muscolatura liscia delle vie respiratorie esercitano nel promuovere la progressione di queste patologie e nel contribuire ai sintomi ad esse associati. L’eccessiva contrazione, infatti, nei pazienti asmatici o affetti da polmonite cronica ostruttiva può impedire completamente gli scambi gassosi e metterne la vita in serio pericolo, mentre i broncodilatatori utilizzati come farmaci di prima scelta possono avere effetti avversi e spesso non sono sufficientemente efficaci.
L’European position paper on rhinosinusitis and nasal polyps del 2012 (EPOS 2012) definisce la rinosinusite nell’adulto e nel bambino come una infiammazione dei seni paranasali, caratterizzata da due o più sintomi, tra i quali ostruzione respiratoria, congestione o secrezione nasale più o meno dolore e pressione facciale, iposmia o tosse nei bambini, in associazione a segni endoscopici e radiologici.
L’interazione microrganismo-ospite nella CRS e, in particolare, le infezioni batteriche e l’immunità innata sono state ampiamente studiate in letteratura al fine di chiarire i meccanismi patogenetici della malattia e di ottimizzare gli interventi terapeutici14. Particolare attenzione è stata dedicata alle CRS refrattarie e recalcitranti.
La prevalenza di CRS in assenza di difetti immunitari sistemici clinicamente evidenti suggerisce un ruolo patogenetico della disfunzione immunitaria innata locale, in particolare nelle rinosinusiti refrattarie e recalcitranti. Tuttavia la risposta immunitaria innata agli agenti microbici e il contributo del signaling epiteliale nella prevenzione delle infezioni a carico delle alte vie respiratorie sono meccanismi complessi e ancora poco conosciuti.
La clearance mucociliare (MCC) rappresenta la principale difesa delle vie respiratorie contro gli agenti patogeni. La frequenza del battito ciliare, infatti, aumenta in risposta a una noxa patogena, come ad esempio alcune componenti batteriche, che, attraverso l’attivazione di secondi messaggeri, determinano un aumento della concentrazione del Ca++ intracellulare e la produzione di NO dotato di una riconosciuta attività antimicrobica.
Sebbene la MCC sia fondamentale nella difesa dell’ospite contro le infezioni batteriche, solo alcune patologie correlate ad alterazioni della funzione ciliare (sindrome di Kartagener) sono chiaramente associate a un aumento delle infezioni respiratorie; in molti altri casi, invece, i meccanismi alla base dell’aumentata suscettibilità alle infezioni respiratorie in generale e alle CRS in particolare non sono ancora stati caratterizzati.
Qual è la funzione del TAS2R38?
La maggior parte degli agenti microbici, come già discusso in precedenza, secerne segnali chimici noti come molecole quorum-sensing, che regolano l’espressione dei geni coinvolti nella formazione del biofilm, nella virulenza e in altri processi vitali per i microrganismi. Tra le molecole quorumsensing l’AHL prodotta da Pseudomonas aeruginosa sarebbe in grado di interagire con i recettori T2Rs per l’amaro presenti nell’epitelio respiratorio.
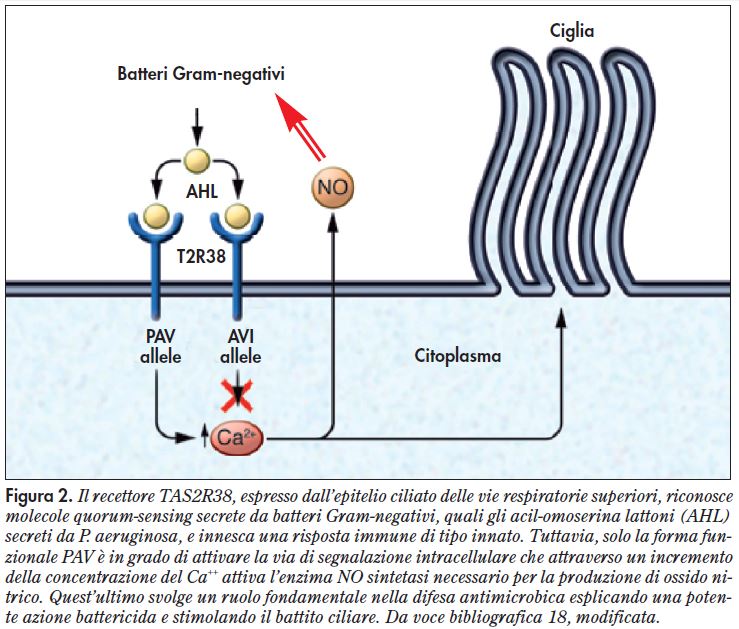
Un recente studio ha dimostrato che le molecole AHLs di P. aeruginosa attivano il recettore per l’amaro T2R38 espresso dalle cellule della mucosa rinosinusale, mentre mutanti di P. aeruginosa privi di AHL non sono in grado di attivarlo. L’attivazione del recettore determina un aumento del flusso di Ca++ e della frequenza del battito ciliare, oltre a stimolare la produzione di NO che agisce come battericida nei confronti del patogeno12.
Quali sono i polimorfismi del TAS2R38?
I polimorfismi del recettore TAS2R38 possono conferire una differente suscettibilità alle infezioni respiratorie
Il TAS2R38 esibisce nella popolazione caucasica tre comuni polimorfismi Pro49Ala, Ala262Val, Val296Ile, che danno luogo ai due principali aplotipi PAV e AVI15. L’espressione dell’uno o dell’altro aplotipo dà luogo rispettivamente alla forma funzionale e a quella non funzionale del recettore, incapace quest’ultima di rispondere a specifici agonisti quali feniltiocarbamide e propiltiouracile. I due aplotipi PAV e AVI segregano in due principali classi fenotipiche: i “percettori”, sensibili all’amaro, sono omo- o eterozigoti per l’allele PAV; i “non percettori”, insensibili, sono omozigoti per l’allele AVI16.
Cosa ci dice TAS2R38 sulle infezioni batteriche?
Studi recenti indicano che le variazioni genetiche del recettore TAS2R38 che condizionano la sensibilità al gusto amaro possono contribuire a determinare le differenze individuali nella suscettibilità alle infezioni delle vie aeree superiori17.
L’ipotesi che la risposta agli AHL possa essere condizionata all’espressione della variante funzionale del recettore è stata esplorata in vitro utilizzando campioni di tessuto naso-sinusale provenienti da pazienti genotipizzati per il recettore T2R38. Lee e coll.12 hanno osservato che nelle colture derivanti da individui omozigoti per l’aplotipo funzionale PAV la produzione di NO, la clearance mucociliare e il potere battericida risultavano di gran lunga più efficaci che nelle colture derivanti da individui omo- o eterozigoti per l’aplotipo non funzionale AVI (Figura 2)18.
In conseguenza di questo risultato era lecito ipotizzare che pazienti con genotipo AVI/PAV o AVI/AVI fossero a maggior rischio di contrarre infezioni da Gram-negativi, rispetto agli omozigoti PAV.
Per determinare se i risultati ottenuti dai test in vitro avessero un significato clinico gli Autori hanno studiato la correlazione tra genotipo TAS2R38 e referti microbiologici sia dei tamponi nasali, sia dei tessuti prelevati in corso di intervento chirurgico per patologie naso-sinusali, tra cui CRS. Il risultato di questa analisi si è rivelato molto interessante, perché ha mostrato una significativa differenza nella frequenza dell’allele non funzionale (AVI) rispetto a quello funzionale (PAV) tra i pazienti le cui colture erano risultate positive a batteri Gram-negativi, inclusa P. aeruginosa.
Infatti, nessuno dei pazienti con infezioni da Gram-negativi o più specificamente da P. aeruginosa era omozigote per l’aplotipo PAV (Tabella I).
In seguito a questi risultati gli stessi Autori hanno effettuato uno studio su pazienti sottoposti a chirurgia endoscopica funzionale dei seni nasali per CRS e/o poliposi nasale, presso l’Università della Pennsylvania (Filadelfia), evidenziando che il polimorfismo TAS2R38 non funzionale (AVI) correla con forme di CRS recalcitrante che non rispondono alla terapia medica e richiedono il trattamento chirurgico molto più frequentemente rispetto al polimorfismo funzionale19. Infatti, nel campione studiato, la frequenza del genotipo PAV/PAV era significativamente più bassa (6%) rispetto alla popolazione generale (20%), mentre il genotipo AVI/AVI era significativamente più frequente (37%) rispetto alla popolazione di riferimento (29%).
Quindi, lo studio suggeriva una importante correlazione clinica tra genotipo TAS2R38, suscettibilità alle infezioni delle vie respiratorie superiori e necessità del trattamento chirurgico nella gestione clinica delle CRS.
Complessivamente i dati che derivano da questi studi suggeriscono interessanti implicazioni cliniche, per esempio la possibilità di predire la suscettibilità alle infezioni batteriche e/o la necessità del ricorso alla chirurgia attraverso un “semplice” test genetico per la determinazione del diplotipo TAS2R38.
Il "sapore" dei microbi
In conclusione, il genotipo TASR38 omozigote PAV, a differenza dei genotipi AVI omo- o eterozigote, sarebbe associato all’assenza di infezioni rinosinusali da Gram-negativi o specificamente da P. aeruginosa e poiché la colonizzazione batterica delle alte vie respiratorie precede e prelude a infezioni delle basse vie respiratorie in soggetti particolarmente predisposti, lo studio dei TAS2Rs potrebbe avere un ruolo cruciale non solo nella prevenzione delle flogosi delle basse vie respiratorie oltre che delle rinosinusiti, ma anche nella programmazione terapeutica di queste ultime, riservando il trattamento chirurgico solo a casi veramente selezionati. Inoltre, lo studio dei meccanismi molecolari alla base della formazione del biofilm può sostenere la ricerca farmacologica nello studio di nuove molecole in grado di prevenirne la formazione e/o determinarne la distruzione. Se poi si pensa che i TAS2Rs potrebbero essere in grado di contribuire anche ai meccanismi di immunità innata, questo campo di ricerca potrebbe aprire delle prospettive terapeutiche innovative e del tutto inaspettate.
Per molti anni l’ACP con Medico e Bambino si è occupata del “bambino catarrale” che trascorre intere stagioni tra un’infezione e un’altra, continuamente contagiato dai compagni di nido e di asilo (indimenticabili interventi del prof. Panizon!). Sì, “benedetto asilo”, perché aiuta a modulare la risposta immune per il futuro adulto, ma quanto disagio per la famiglia e il pediatra! Abbiamo per anni cercato i marcatori della suscettibilità individuale: per 20 anni si è puntato sulle capacità di risposta immune dell’individuo, ma non sono mai state evidenziate anomalie significative. Eppure ci devono essere dei fattori di suscettibilità che distinguono bambini sempre malati da bambini che vanno avanti indenni nello stesso ambiente, e spesso nella stessa famiglia. Certamente polimorfismi genici con significato funzionale dovranno essere coinvolti nella differente suscettibilità di diversi bambini: il campo è ancora tutto da esplorare.
Bibliografia
- Morini G, Greco L. Lo sviluppo del gusto nel bambino. Medico e Bambino 2010;29(7): 509-13.
- Di Salle F, Cantone E, Savarese MF, et al. Effect of Carbonation on Brain Processing of Sweet Stimuli in Humans. Gastroenterology 2013;145(3):537-9.
- Bachmanov A, Beauchamp G. Taste receptor genes. Annu Rev Nutr 2007;27:389-414.
- Negri R, Morini G, Greco L. From the tongue to the gut. J Pediatr Gastroenterol Nutr 2011;53: 601-5.
- Behrens M, Meyerhof W. Gustatory and extragustatory functions of mammalian taste receptors. Physiol Behav 2011;105:4-13.
- Gerspach AC, Steinert RE, Schonenberger L, Graber-Maier A, Beglinger C. The role of the gut sweet taste receptor in regulating GLP-1, PYY and CKK release in humans. Am J Physiol Endocrinol Metab 2011;301(2):E317-25.
- Rozengurt E, Sternini C. Taste receptor signaling in the mammalian gut. Curr Opin Pharmacol 2007;7(6):557-62.
- Tizzano M, Gulbransen BD, Vandenbeuch A, et al. Nasal chemosensory cells use bitter taste signaling to detect irritants and bacterial signals. Proc Natl Acad Sci USA 2010;107: 3210-5.
- Finger TE, Böttger B, Hansen A, Anderson KT, Alimohammadi H, Silver WL. Solitary chemoreceptor cells in the nasal cavity serve as sentinels of respiration. Proc Natl Acad Sci USA 2003;100(15):8981-6.
- Sbarbati A, Tizzano M, Merigo F, et al. Acyl homoserine lactones induce early response in the airway. Anat Rec (Hoboken) 2009;292(3): 439-48.
- Shah AS, Ben-Shahar Y, Moninger TO, Kline JN, Welsh MJ. Motile cilia of human airway epithelia are chemosensory. Science 2009;325: 1131-4.
- Lee RJ, Xiong G, Kofonow JM, et al. T2R38 taste receptor polymorphisms underlie susceptibility to upper respiratory infection. J Clin Invest
- 2012;122:4145-59.
- Zhang CH, Lifshitz LM, Uy KF, Ikebe M, Fogarty KE, ZhuGe R. The cellular and molecular basis of bitter tastant-induced bronchodilation. PLoS Biol 2013;11(3):e1001501.
- Hamilos DL. Host-microbial interactions in patients with chronic rhinosinusitis. J Allergy Clin Immunol 2013;131(4):1263-4.
- Kim UK, Jorgenson E, Coon H, Leppert M, Risch N, Drayna D. Positional cloning of the human quantitative trait locus underlying taste sensitivity to phenylthiocarbamide. Science 2003;299:1221-5.
- Bufe B, Breslin PA, Kuhn C, et al. The molecular basis of individual differences in phenylthiocarbamide and propylthiouracil bitterness perception. Curr Biol 2005;15:322-7.
- Lee RJ, Cohen NA. The emerging role of the bitter taste receptor T2R38 in upper respiratory infection and chronic rhinosinusitis. Am J Rhinol Allergy 2013;27(4):283-6.
- Prince A. The bitter taste of infection. J Clin Invest 2012;122(11):3847-9.
- Adappa ND, Howland TJ, Palmer JN, et al. Genetics of the taste receptor T2R38 correlates with chronic rhinosinusitis necessitating surgical intervention. Int Forum Allergy Rhinol 2013; 3(3):184-7.

